Gametes Can Only Carry One Allele for Each Inherited Characteristic.
Genes come in unlike varieties, chosen alleles. Somatic cells contain 2 alleles for every gene, with one allele provided by each parent of an organism. Oftentimes, it is incommunicable to determine which ii alleles of a factor are nowadays within an organism's chromosomes based solely on the outward appearance of that organism. Withal, an allele that is hidden, or not expressed by an organism, tin still exist passed on to that organism's offspring and expressed in a later generation.
Tracing a subconscious cistron through a family unit tree
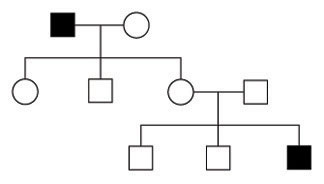
Figure 1: In this family full-blooded, black squares point the presence of a detail trait in a male, and white squares represent males without the trait. White circles are females. A trait in 1 generation tin be inherited, only not outwardly apparent before two more than generations (compare black squares).
The family tree in Figure 1 shows how an allele tin disappear or "hibernate" in i generation and then reemerge in a later generation. In this family tree, the begetter in the beginning generation shows a particular trait (as indicated by the blackness foursquare), just none of the children in the second generation show that trait. Still, the trait reappears in the third generation (blackness square, lower correct). How is this possible? This question is best answered by considering the bones principles of inheritance.
Mendel'southward principles of inheritance
Gregor Mendel was the showtime person to describe the manner in which traits are passed on from i generation to the side by side (and sometimes skip generations). Through his breeding experiments with pea plants, Mendel established three principles of inheritance that described the transmission of genetic traits before genes were even discovered. Mendel'south insights greatly expanded scientists' understanding of genetic inheritance, and they as well led to the evolution of new experimental methods.
One of the key conclusions Mendel reached after studying and breeding multiple generations of pea plants was the idea that "[you cannot] draw from the external resemblances [whatsoever] conclusions equally to [the plants'] internal nature." Today, scientists use the word "phenotype" to refer to what Mendel termed an organism's "external resemblance," and the word "genotype" to refer to what Mendel termed an organism'due south "internal nature." Thus, to restate Mendel'due south decision in modern terms, an organism's genotype cannot be inferred by simply observing its phenotype. Indeed, Mendel's experiments revealed that phenotypes could be hidden in i generation, only to reemerge in subsequent generations. Mendel thus wondered how organisms preserved the "elementen" (or hereditary cloth) associated with these traits in the intervening generation, when the traits were hidden from view.
How do subconscious genes pass from i generation to the next?
Although an individual gene may code for a specific concrete trait, that factor can exist in different forms, or alleles. One allele for every gene in an organism is inherited from each of that organism'due south parents. In some cases, both parents provide the aforementioned allele of a given gene, and the offspring is referred to as homozygous ("man" significant "aforementioned") for that allele. In other cases, each parent provides a different allele of a given cistron, and the offspring is referred to as heterozygous ("hetero" meaning "different") for that allele. Alleles produce phenotypes (or physical versions of a trait) that are either ascendant or recessive. The dominance or recessivity associated with a particular allele is the result of masking, by which a dominant phenotype hides a recessive phenotype. By this logic, in heterozygous offspring simply the dominant phenotype volition be credible.
The human relationship of alleles to phenotype: an example
Relationships betwixt ascendant and recessive phenotypes can exist observed with convenance experiments. Gregor Mendel bred generations of pea plants, and as a result of his experiments, he was able to suggest the idea of allelic cistron forms. Modern scientists employ organisms that have faster breeding times than the pea plant, such as the fruit fly (Drosophila melanogaster). Thus, Mendel'southward principal discoveries will be described in terms of this modern experimental choice for the rest of this give-and-take.

Figure 2: In fruit flies, two possible body color phenotypes are chocolate-brown and black.
The substance that Mendel referred to equally "elementen" is now known as the gene, and different alleles of a given gene are known to give rise to different traits. For instance, breeding experiments with fruit flies have revealed that a single gene controls fly body colour, and that a fruit fly can have either a brown body or a black torso. This coloration is a direct result of the body colour alleles that a wing inherits from its parents (Figure 2).
In fruit flies, the factor for body color has two different alleles: the blackness allele and the brown allele. Moreover, brown body color is the ascendant phenotype, and black torso color is the recessive phenotype.
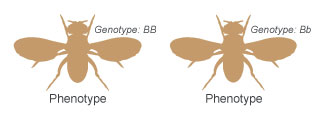
Figure 3: Dissimilar genotypes tin produce the same phenotype.
Researchers rely on a type of autograph to stand for the dissimilar alleles of a cistron. In the case of the fruit fly, the allele that codes for brown torso color is represented past a B (because brown is the dominant phenotype), and the allele that codes for black body color is represented by a b (because blackness is the recessive phenotype). Equally previously mentioned, each wing inherits ane allele for the body colour factor from each of its parents. Therefore, each fly will carry two alleles for the body color factor. Within an individual organism, the specific combination of alleles for a gene is known equally the genotype of the organism, and (as mentioned above) the physical trait associated with that genotype is called the phenotype of the organism. So, if a fly has the BB or Bb genotype, it volition accept a brown body color phenotype (Figure 3). In contrast, if a wing has the bb genotype, it will have a black body phenotype.
Dominance, breeding experiments, and Punnett squares
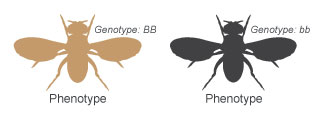
Figure 4: A brown fly and a black fly are mated.
The best way to understand the potency and recessivity of phenotypes is through breeding experiments. Consider, for instance, a breeding experiment in which a fruit wing with brownish body color (BB) is mated to a fruit wing with black body colour (bb). (The genotypes of these two flies are shown in Effigy 4.) The breeding, or cross, performed in this experiment can be denoted as BB × bb.

Figure 5: A Punnett foursquare.
When conducting a cantankerous, one manner of showing the potential combinations of parental alleles in the offspring is to align the alleles in a grid called a Punnett square, which functions in a manner similar to a multiplication table (Figure v).
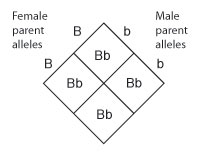
Figure half dozen: Each parent contributes one allele to each of its offspring. Thus, in this cross, all offspring volition take the Bb genotype.
If the alleles on the outside of the Punnett square are paired up in each intersecting square in the grid, it becomes clear that, in this particular cross, the female parent can contribute but the B allele, and the male parent can contribute only the b allele. Every bit a result, all of the offspring from this cross will have the Bb genotype (Effigy 6).
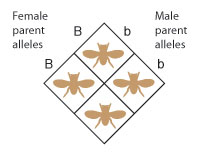
Figure 7: Genotype is translated into phenotype. In this cross, all offspring will take the chocolate-brown body color phenotype.
If these genotypes are translated into their corresponding phenotypes, all of the offspring from this cantankerous will take the brown trunk color phenotype (Effigy 7).
This effect shows that the brown allele (B) and its associated phenotype are dominant to the black allele (b) and its associated phenotype. Even though all of the offspring take brown torso color, they are heterozygous for the black allele.
The phenomenon of ascendant phenotypes arising from the allele interactions exhibited in this cross is known as the principle of uniformity, which states that all of the offspring from a cross where the parents differ past just one trait volition appear identical.
How can a convenance experiment exist used to detect a genotype?

Effigy viii: A Punnett square can help determine the identity of an unknown allele.
Brown flies tin be either homozygous (BB) or heterozygous (Bb) - but is it possible to make up one's mind whether a female fly with a chocolate-brown body has the genotype BB or Bb? To answer this question, an experiment chosen a test cross can be performed. Test crosses help researchers determine the genotype of an organism when only its phenotype (i.e., its advent) is known.
A test cross is a breeding experiment in which an organism with an unknown genotype associated with the dominant phenotype is mated to an organism that is homozygous for the recessive phenotype. The Punnett square in Figure 8 can be used to consider how the identity of the unknown allele is adamant in a test cross.
Convenance the flies shown in this Punnett square will decide the distribution of phenotypes amidst their offspring. If the female parent has the genotype BB, all of the offspring volition have brownish bodies (Figure ix, Issue 1). If the mother has the genotype Bb, l% of the offspring will have brown bodies and 50% of the offspring will have black bodies (Effigy ix, Consequence 2). In this manner, the genotype of the unknown parent tin can be inferred.

Again, the Punnett squares in this example function like a genetic multiplication table, and there is a specific reason why squares such as these work. During meiosis, chromosome pairs are split apart and distributed into cells called gametes. Each gamete contains a unmarried copy of every chromosome, and each chromosome contains 1 allele for every gene. Therefore, each allele for a given gene is packaged into a separate gamete. For example, a fly with the genotype Bb will produce two types of gametes: B and b. In comparing, a fly with the genotype BB volition just produce B gametes, and a wing with the genotype bb volition merely produce b gametes.

Figure 10: A monohybrid cross betwixt two parents with the Bb genotype.
The following monohybrid cantankerous shows how this concept works. In this type of convenance experiment, each parent is heterozygous for body colour, so the cross can be represented by the expression Bb × Bb (Effigy x).
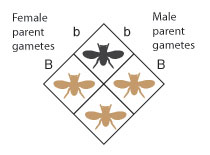
Figure 11: The phenotypic ratio is iii:1 (brown body: black body).
The outcome of this cantankerous is a phenotypic ratio of iii:1 for brown body color to black torso colour (Effigy 11).
This observation forms the second principle of inheritance, the principle of segregation, which states that the two alleles for each gene are physically segregated when they are packaged into gametes, and each parent randomly contributes one allele for each gene to its offspring.
Tin two different genes be examined at the aforementioned time?
The principle of segregation explains how individual alleles are separated amid chromosomes. But is information technology possible to consider how ii different genes, each with different allelic forms, are inherited at the same time? For example, can the alleles for the body color gene (brownish and blackness) be mixed and matched in different combinations with the alleles for the eye color gene (ruby-red and brown)?
The simple answer to this question is aye. When chromosome pairs randomly align along the metaphase plate during meiosis I, each member of the chromosome pair contains one allele for every factor. Each gamete will receive 1 copy of each chromosome and one allele for every gene. When the individual chromosomes are distributed into gametes, the alleles of the different genes they acquit are mixed and matched with respect to one another.
In this example, there are two different alleles for the eye colour cistron: the E allele for ruby eye colour, and the e allele for brown middle colour. The reddish (Due east) phenotype is dominant to the dark-brown (e) phenotype, and so heterozygous flies with the genotype Ee will have red eyes.

Effigy 12: The four phenotypes that can result from combining alleles B, b, E, and e.
When two flies that are heterozygous for chocolate-brown body color and cherry eyes are crossed (BbEe X BbEe), their alleles can combine to produce offspring with four different phenotypes (Figure 12). Those phenotypes are chocolate-brown torso with red optics, brown body with dark-brown eyes, black body with blood-red optics, and black torso with chocolate-brown optics.
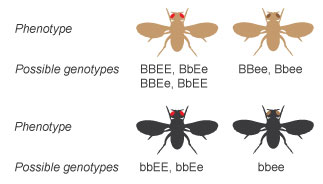
Figure 13: The possible genotypes for each of the four phenotypes.
Fifty-fifty though but four different phenotypes are possible from this cantankerous, ix unlike genotypes are possible, as shown in Figure 13.
The dihybrid cross: charting ii different traits in a unmarried breeding experiment
Consider a cross betwixt ii parents that are heterozygous for both body color and eye color (BbEe x BbEe). This type of experiment is known every bit a dihybrid cross. All possible genotypes and associated phenotypes in this kind of cantankerous are shown in Figure xiv.
The four possible phenotypes from this cantankerous occur in the proportions 9:three:3:1. Specifically, this cross yields the post-obit:
- ix flies with brown bodies and red eyes
- 3 flies with brownish bodies and brown eyes
- iii flies with black bodies and red eyes
- i fly with a black body and brown optics
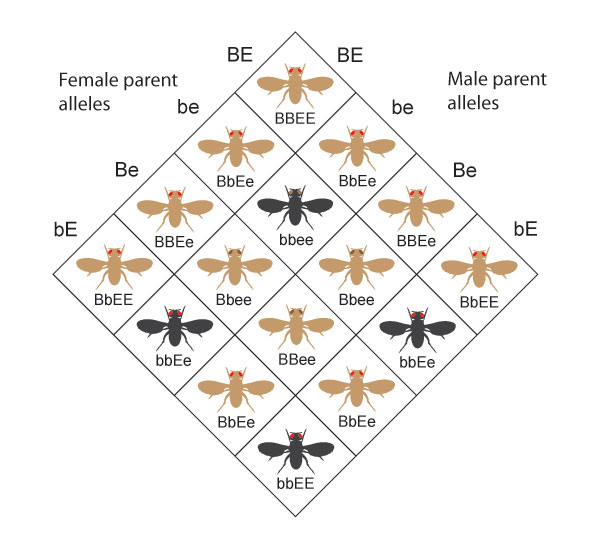
Figure 14: These are all of the possible genotypes and phenotypes that can result from a dihybrid cantankerous between two BbEe parents.
Why does this ratio of phenotypes occur? To answer this question, information technology is necessary to consider the proportions of the individual alleles involved in the cross. The ratio of brown-bodied flies to black-bodied flies is 3:1, and the ratio of cerise-eyed flies to brown-eyed flies is also iii:ane. This ways that the outcomes of body colour and eye color traits appear every bit if they were derived from two parallel monohybrid crosses. In other words, fifty-fifty though alleles of two different genes were involved in this cross, these alleles behaved equally if they had segregated independently.
The issue of a dihybrid cross illustrates the third and final principle of inheritance, the master of independent assortment, which states that the alleles for one cistron segregate into gametes independently of the alleles for other genes. To restate this principle using the example above, all alleles assort in the same way whether they code for body colour solitary, middle colour alone, or both body color and heart color in the same cross.
The impact of Mendel'south principles
Seminal experiments on inheritance
Mendel'due south principles can exist used to understand how genes and their alleles are passed down from one generation to the side by side. When visualized with a Punnett square, these principles can predict the potential combinations of offspring from 2 parents of known genotype, or infer an unknown parental genotype from tallying the resultant offspring.
An important question still remains: Do all organisms pass on their genes in this way? The answer to this question is no, but many organisms exercise exhibit unproblematic inheritance patterns similar to those of fruit flies and Mendel's peas. These principles class a model against which different inheritance patterns can be compared, and this model provide researchers with a way to analyze deviations from Mendelian principles.
Source: http://www.nature.com/scitable/topicpage/inheritance-of-traits-by-offspring-follows-predictable-6524925
0 Response to "Gametes Can Only Carry One Allele for Each Inherited Characteristic."
Post a Comment